












Heermann's Gull Larus heermanni Scientific name definitions
- NT Near Threatened
- Names (28)
- Monotypic
Kamal Islam and Enriqueta Velarde
Version: 2.0 — Published April 9, 2020
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Bulgarian | Сивокорема чайка |
| Catalan | gavina de Heermann |
| Croatian | kalifornijski galeb |
| Czech | racek mexický |
| Danish | Askemåge |
| Dutch | Heermannmeeuw |
| English | Heermann's Gull |
| English (United States) | Heermann's Gull |
| Estonian | tuhkkajakas |
| Finnish | meksikonlokki |
| French | Goéland de Heermann |
| French (Canada) | Goéland de Heermann |
| German | Heermannmöwe |
| Icelandic | Hvinmáfur |
| Japanese | オグロカモメ |
| Norwegian | askemåke |
| Polish | mewa śniada |
| Russian | Серобрюхая чайка |
| Serbian | Hirmanov galeb |
| Slovak | čajka pobrežná |
| Slovenian | Heermannov galeb |
| Spanish | Gaviota Mexicana |
| Spanish (Costa Rica) | Gaviota de Heermann |
| Spanish (Mexico) | Gaviota Plomiza |
| Spanish (Spain) | Gaviota mexicana |
| Swedish | vithuvad mås |
| Turkish | Heermann Martısı |
| Ukrainian | Мартин червонодзьобий |
Larus heermanni Cassin, 1852
PROTONYM:
Larus Heermanni
Cassin, 1852. Proceedings of the Academy of Natural Sciences of Philadelphia 6, p.187.
TYPE LOCALITY:
San Diego, California.
SOURCE:
Avibase, 2024
Definitions
- LARUS
- heermani / heermanni
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Content Partner

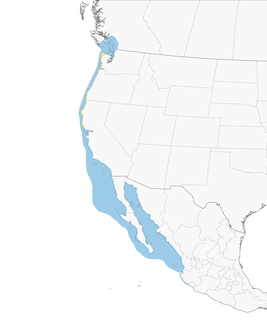
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Heermann's Gull


































