












Black Stork Ciconia nigra Scientific name definitions
- LC Least Concern
- Names (63)
- Monotypic
Andrew Elliott, David Christie, Ernest Garcia, and Peter F. D. Boesman
Version: 1.0 — Published March 4, 2020
Text last updated November 30, 2018
Text last updated November 30, 2018
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Grootswartooievaar |
| Albanian | Lejleku i zi |
| Arabic | لقلق اسود |
| Armenian | Սև արագիլ |
| Assamese | কালশৰ |
| Asturian | Cigüeña prieta |
| Azerbaijani | Qara leylək |
| Bangla (India) | কালোজঙ্ঘা |
| Basque | Zikoina beltza |
| Bulgarian | Черен щъркел |
| Catalan | cigonya negra |
| Chinese | 黑鸛 |
| Chinese (Hong Kong SAR China) | 黑鸛 |
| Chinese (SIM) | 黑鹳 |
| Croatian | crna roda |
| Czech | čáp černý |
| Danish | Sort Stork |
| Dutch | Zwarte ooievaar |
| English | Black Stork |
| English (United States) | Black Stork |
| Estonian | must-toonekurg |
| Finnish | mustahaikara |
| French | Cigogne noire |
| French (Canada) | Cigogne noire |
| Galician | Cegoña negra |
| Georgian | შავი ყარყატი (იშხვარი) |
| German | Schwarzstorch |
| Greek | Μαύρος Πελαργός |
| Gujarati | કાળો ઢોંક |
| Hebrew | חסידה שחורה |
| Hindi | सुरमल |
| Hungarian | Fekete gólya |
| Icelandic | Kolstorkur |
| Italian | Cicogna nera |
| Japanese | ナベコウ |
| Kannada | ಕರಿ ಕೊಕ್ಕರೆ |
| Korean | 먹황새 |
| Ladakhi | གཞད་ནག |
| Latvian | Melnais stārķis |
| Lithuanian | Juodasis gandras |
| Malayalam | കരിമ്പകം |
| Mongolian | Хар өрөвтас |
| Nepali (India) | कालो सारस |
| Nepali (Nepal) | कालो गरुड |
| Norwegian | svartstork |
| Persian | لک لک سیاه |
| Polish | bocian czarny |
| Portuguese (Angola) | Cegonha-preta |
| Portuguese (Portugal) | Cegonha-preta |
| Punjabi (India) | ਕਾਲਾ ਲਮਢੀਂਗ |
| Romanian | Barză neagră |
| Russian | Чёрный аист |
| Serbian | Crna roda |
| Slovak | bocian čierny |
| Slovenian | Črna štorklja |
| Spanish | Cigüeña Negra |
| Spanish (Spain) | Cigüeña negra |
| Swedish | svart stork |
| Telugu | నల్ల కొంగ |
| Thai | นกกระสาดำ |
| Turkish | Kara Leylek |
| Ukrainian | Лелека чорний |
| Zulu | unowanga |
Ciconia nigra (Linnaeus, 1758)
PROTONYM:
Ardea nigra
Linnaeus, 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decima, reformata 1, p.142.
TYPE LOCALITY:
Northern Europe; restricted type locality, Sweden.
SOURCE:
Avibase, 2024
Definitions
- CICONIA
- ciconia
- nigra
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Originally Appeared in

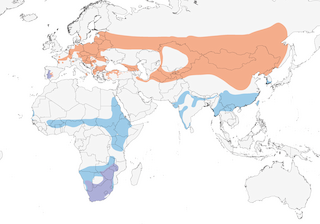
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Black Stork



































