







Buff-throated Monal-Partridge Tetraophasis szechenyii Scientific name definitions
- LC Least Concern
- Names (24)
- Monotypic
Philip J. K. McGowan, Guy M. Kirwan, and Eduardo de Juana
Version: 1.0 — Published March 4, 2020
Text last updated May 23, 2017
Text last updated May 23, 2017
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Bulgarian | Оранжевогуш монал |
| Catalan | gall nival de Szecheny |
| Chinese (SIM) | 黄喉雉鹑 (四川雉鹑) |
| Croatian | pjegavoboka jarebica |
| Czech | velekur plavohrdlý |
| Dutch | Roodkeelwigstaarthoen |
| English | Buff-throated Monal-Partridge |
| English (United States) | Buff-throated Monal-Partridge |
| Estonian | koldkurk-mägikana |
| Finnish | metsälumipyy |
| French | Tétraophase de Széchenyi |
| French (Canada) | Tétraophase de Széchenyi |
| German | Rostkehl-Keilschwanzhuhn |
| Japanese | チベットキジシャコ |
| Norwegian | gulstrupehøne |
| Polish | górak płowogardły |
| Russian | Светлогорлый кундык |
| Serbian | Žutobrada fazanska jarebica |
| Slovak | tokáň čečinový |
| Spanish | Perdiz Faisán Gorjiclara |
| Spanish (Spain) | Perdiz faisán gorjiclara |
| Swedish | gulstrupig kungshöna |
| Turkish | Sarı Gerdanlı Kumar Kekliği |
| Ukrainian | Кундик жовтогорлий |
Tetraophasis szechenyii Madarász, 1885
PROTONYM:
Tetraophasis Széchenyii
Madarász, 1885. Zeitschrift für die gesammte Ornithologie 2, p.50 pl.2.
TYPE LOCALITY:
'Szii-csuan ' = Szechuan or eastern Tibet.
SOURCE:
Avibase, 2024
Definitions
- TETRAOPHASIS
- szechenyii
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Originally Appeared in

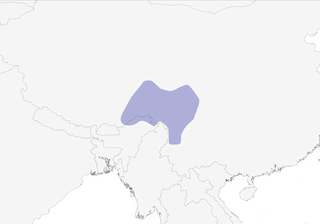
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Buff-throated Monal-Partridge



































